


Sadales vārpsta ir pagaidu struktūraveidojas mitozes un meiozes procesos, nodrošinot hromosomu un šūnu dalīšanās segregāciju. Tas ir bipolārs: mikrotubulu sistēma, kas veidota telpās starp stabiem, atgādina formas vārpstu. Centromēra reģionā vārpstas mikrotubulas pievienojas hromosomu kinetokorām. Uz tiem kromosomas pārvietojas uz stabiem.

Sadales vārpsta sastāv no trim galvenajiemstrukturālie elementi: mikrotubulas, sadalīšanas stabi un hromosomas. Sadalīšanas stabi dzīvniekiem tiek organizēti ar centrosomu palīdzību, kas satur centrioles. Gadījumā, ja nav centrosomu (augos un oocītos dažās dzīvnieku sugās), vārpstai ir plaši stabi un to sauc par acentrosomālu. Vēl viena struktūra ir saistīta ar vārpstas - motoru proteīnu veidošanos. Viņi pieder dyneinam un kinesinam.
Vārpsta ir bipolāra struktūra.Abos stabos atrodas centrosomas - organellas, kas ir mikrotubulāru organizācijas centri. Centrosomas struktūrā ir divi centrioli, kurus ieskauj daudzi dažādi proteīni. Starp poliem atrodas kondensēti hromosomi, kam ir divi hromatīdi, kas pievienoti centromēra vietai. Centromēras reģionā ir kinetokori, kuriem ir pievienoti mikrotubulas.
Tā kā sadalīšanas vārpsta ir struktūra,par šūnu dalīšanos, tās montāžas sākums notiek propāzē. Augos un oocītos, ja nav centrosomu, kodola apvalks kalpo par mikrotubulu organizācijas centru. Mikrotubulas vēršas pie kodolmateriāla aploksnes un propāzes fāzes beigās to orientācija beidzas, un tiek veidots "propāzes ass" - nākamās sadalītāja vārpstas ass.
Sakarā ar to, ka dzīvnieku šūnāsCentrosoms kalpo kā organizācijas centrs, sadalīšanas vārpstas veidošanās sākums ir divu centrosomu novirze prophāzes periodā. Tas ir iespējams, pateicoties motora proteīniem dyneinam: tie ir piestiprināti pie kodola ārējās virsmas, kā arī šūnu membrānas iekšpusē. Delmīna grupa, kas piestiprināta pie membrānas, savienojas ar astrālajiem mikrotubuliem un sāk kustēties pret mīnusa galu, kuru dēļ centrosomas atšķaida pretējā šūnu membrānas daļā.
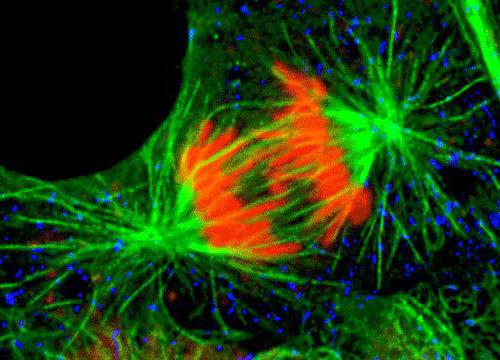
Vārpstas sadalījuma galīgais veidojumsnotiek prometafāzes stadijā, pēc kodola membrānas pazušanas, tā kļūst pilnvērtīga, jo pēc tam centrosome un mikrotubulas var piekļūt vārpstas komponentiem.
Tomēr ir viens izņēmums: jaunos raugos kodola sadalīšanās vārpstas veidošanās notiek kodola iekšienē.
Kvēldiega vārpstas un to veidošanāsorientācija ir neiespējama bez diviem procesiem: mikrotubulu organizēšana ap hromosomām un to piesaiste viens otram pretējā sadalījuma stabos. Daudzi no elementiem, kas nepieciešami sadalīšanas vārpstas galīgai veidošanai, ieskaitot hromosomas un motoriskās olbaltumvielas, atrodas šūnu kodolā, un mikrotubulas, un, ja tā ir dzīvnieku šūna, centrosomi atrodas citoplazmā, ti, komponenti ir izolēti viens no otra. Tāpēc vārpstas veidošanās beidzas tikai pēc kodolkoka izzušanas.

Sadalīšanās vārpstas veidošanā ir iesaistīts proteīnsir arī daudzas citas struktūras, un šis process ir labi pētīts dzīvnieku šūnās. Prophāzes periodā mikrotubulas veido centrālās formas staru struktūru, kas atšķiras radiālā virzienā. Pēc kodola membrānas iznīcināšanas dinamiski nestabilas mikrotubulas sāk aktīvi pārbaudīt šo zonu, un uz tām var noteikt hromosomu kinetokorus. Dažas hromosomas uzreiz parādās pretējos stabos, bet pārējie vispirms ir saistīti ar viena pola mikrotubulām, un tikai tad sāk virzīties uz vēlamo polu. Kad process ir pabeigts, hromosomas, kas jau ir saistītas ar polu, sāk pievienoties mikrotubulām no pretējā poliņa ar kinetokoriem, tādējādi no desmit līdz četrdesmit mēģenēm metafāzes procesa laikā ir pievienotas kinetokorām. Šo veidošanos sauc par kinetohorny gaismu. Pakāpeniski katrs hromosomos ir saistīts ar pretējo polu, un tie veido metafāzes plati sadalīšanas vārpstas centrālajā daļā.

Ir vēl viens scenārijs, kurāveidojas vārpsta. Tas ir iespējams šūnām ar centrosomām un šūnām, kurās tās nav. Procesā ir iesaistīts gamma-tubulīna gredzena komplekss, tādēļ īsu mikrotubulu kodācija notiek hromosomu tuvumā. Caurules ir pievienotas kinetokoriem ar plus-end, pēc kura sākas mikrotubulu polimerizācija, ti, kontrolēta augšana. Negatīvie gali "apvienojas" un paliek pie sadales stabiem, kas saistīti ar motoru proteīniem. Ja sadalīšanas vārpstas veidošanā ir iesaistīts centrosomas pāris, tas atvieglo mikrotubulu savienošanu, bet process ir iespējams bez tiem.

Skaidra hromosomu atdalīšana starp divām šūnāmveidojas sadalīšanas laikā, var notikt tikai tad, ja pāris hromatīdi ar to kinetokoriem pievienojās dažādiem stabiem. Kromatīdu bipolārā neatbilstība tiek saukta par amfitepisko, bet ir arī citas iespējas, kas rodas, sadalot vārpstu. Tas ir monotheisks (viens kinetochore ir savienots ar vienu un to pašu pole) un sintētisks (abi hromosomas kinetokori ir savienoti ar vienu polu). Merothepiskā gadījumā kinetochore uzreiz uzņem divus stabus. Tikai normāla, bipolāra sajaukšanās, kas notiek polu spriedzes spēku dēļ, ir stabila, citas savienošanas metodes ir nestabilas un atgriezeniskas, bet ir iespējamas kinetokoriju atrašanās vietas dēļ.


