


Растительный мир - одно из главных богатств нашей כדור הארץ. בזכות הצומח בכדור הארץ יש חמצן שכולנו נושמים, יש בסיס מזון עצום עליו כל החיים חיים תלויים. צמחים הם ייחודיים בכך שהם יכולים להמיר תרכובות כימיות בעלות אופי אורגני לחומרים אורגניים.
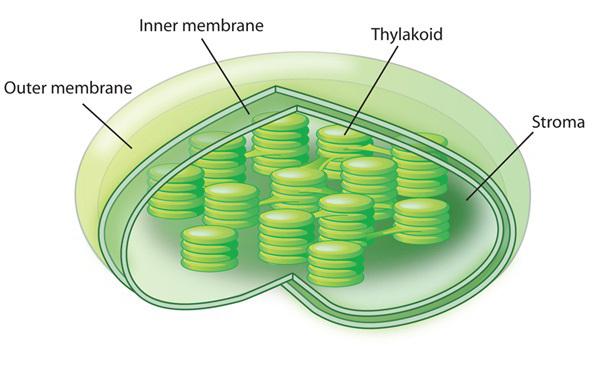
Так называются специфические структуры, в которых מתרחשים תהליכי פוטוסינתזה אשר מכוונים לכריכה של פחמן דו חמצני ולהיווצרות פחמימות מסוימות. תוצר לוואי הוא חמצן. מדובר באורגנואידים מוארכים, המגיעים לרוחב של 2-4 מיקרון, אורכם מגיע ל 5-10 מיקרון. בכמה מינים של אצות ירוקות ניתן למצוא לפעמים כלורופלסטים ענקיים המוארכים ב 50 מיקרון!
לאותן האצות יכול להיות שונהתכונה: לכל התא יש רק אורגנואיד אחד מסוג זה. בתאים של צמחים גבוהים יותר, לרוב ישנם בטווח של 10-30 כלורופלסטים. עם זאת, במקרה שלהם עשויים להיות חריגים בהירים. אז, ברקמת העצבית של שאג רגיל, יש 1000 כלורופלסטים לתא. בשביל מה מיועדות כלורופלסטים אלה? הפוטוסינתזה היא העיקרית שלהם, אך רחוקה מהתפקיד היחיד. כדי להבין בבירור את משמעותם בחיי צמח, חשוב לדעת היבטים רבים של מקורם והתפתחותם. כל זה מתואר בחלקו הבא של המאמר.
אז, מה זה כלורופלסט, למדנו.מאיפה הגיע האורגנואידים האלה? איך קרה שלצמחים יש מכשיר כה ייחודי שהופך את הפחמן הדו-חמצני והמים לתרכובות אורגניות מורכבות?
נכון לעכשיו, הנקודה שולטת בקרב מדעניםמבט למוצא האנדוסימביוטי של אורגנואידים אלה, מכיוון שההופעה העצמאית שלהם בתאי צמח מסופקת למדי. ידוע היטב כי חזזית היא סימביוזה של אצות ופטריות. במקביל, אצות חד-תאיות חיות בתוך התא הפטרייתי. כעת מדענים מציעים כי עם הזמן ציאנו-בקטריות פוטוסינתטיות של פעם חדרו לתאי צמחים, ואז איבדו חלקית את "עצמאותן", והעבירו את רוב הגנום לגרעין.

ההשערה האחרונה יחסית של פרוקריוטיםמקורם של אלמנטים אלה לא היה פופולרי במיוחד בקהילה המדעית, רבים חשבו שהוא "בדים של חובבנים". אולם לאחר ניתוח מעמיק של רצפי נוקליאוטידים ב- DNA של כלורופלסט, ההנחה הזו אושרה בצורה מבריקה. התברר כי מבנים אלה דומים ביותר, אפילו קשורים, ל- DNA של תאים חיידקיים. אז, רצף דומה נמצא בציאנובקטריה שחיה חופשית. בפרט, הגנים של מתחם סינתזת ATP התבררו כדומים מאוד, כמו גם ב"מנגנונים "לתמלול ותרגום.
מקדמים שקובעים את תחילת הקריאהמידע גנטי עם DNA, כמו גם רצפי נוקליאוטידים סופניים שאחראים לסיומו, מאורגנים גם הם בדמותם של חיידקים. כמובן שמיליארדי שנים של טרנספורמציות אבולוציוניות הצליחו לבצע שינויים רבים בכלורופלסט, אך הרצפים בגני הכלורופלסט נותרו זהים לחלוטין. וזו הוכחה מוחלטת, מוחלטת, שלכלורופלסטים הייתה פעם אב קדמון פרוקריוטי. אולי זה היה אורגניזם שממנו נבעו גם ציאנובקטריה מודרנית.
אורגנואיד "מבוגר" מתפתח מפרופלאסטיד.זהו אורגנל קטן וחסר צבע לחלוטין, שיש בו רק כמה מיקרונים. הוא מוקף בקרום דו-שכבתי צפוף המכיל DNA מעגלי הספציפי לכלורופלסט. ל"אבות אבות "אלה של האורגנואידים אין מערכת ממברנה פנימית. בשל גודלם הקטן ביותר, המחקר שלהם קשה ביותר, ולכן ישנם מעט מאוד נתונים על התפתחותם.
ידוע שכמה פרוטופלסטידים כאלהקיים בגרעין של כל תא ביצה של בעלי חיים וצמחים. במהלך התפתחות העובר הם מתחלקים ומועברים לתאים אחרים. קל לאמת את זה: תכונות גנטיות הקשורות איכשהו לפלסטידים מועברות רק דרך הקו האימהי.
קרום פנימי של פרוטופלסטידים לאורך זמןהתפתחות בולטת לאורגנואיד. ממבנים אלה צומחות הממברנות התילאקואיד, האחראיות ליצירת גרגירים ולמלות של סטרומה אורגנואיד. בחשיכה מוחלטת, פרוטופסטיד מתחיל להפוך למבשר כלורופלסט (אטיופלסט). אורגנואיד ראשוני זה מאופיין בכך שמבנה גביש מורכב למדי ממוקם בתוכו. ברגע שאור פוגע בעלה של הצמח, הוא נהרס לחלוטין. לאחר מכן נוצר המבנה הפנימי ה"מסורתי "של הכלורופלסט, שנוצר בדיוק על ידי תילאקואידים ולמלות.
כל תא meristemal מכילכמה פרופלסטידים כאלה (מספרם משתנה בהתאם לסוג הצמח וגורמים אחרים). ברגע שרקמה ראשונית זו מתחילה להפוך לעלה, מבשרי האברונים הופכים לכלורופלסטים. לפיכך, עלי חיטה צעירים שהשלימו את צמיחתם כוללים כלורופלסטים בכמות 100-150 חתיכות. המצב קצת יותר מסובך ביחס לאותם צמחים שמסוגלים להצטבר עמילן.

גילינו מהו כלורופלסט, וחושף בו זמניתהקשר של אורגנואיד זה עם מבנים של אורגניזמים פרוקריוטים. כאן המצב דומה: מדענים גילו זה מכבר כי עמילופלסטים, כמו כלורופלסטים, מכילים בדיוק אותו DNA ונוצרים מאותם פרוטופלסטים בדיוק. לכן יש לשקול אותם באותו היבט. למעשה, יש להחשיב עמילופלסטים כסוג מיוחד של כלורופלסט.
ניתן לצייר אנלוגיה בין פרוטופלסטידים לביןתאי גזע. במילים פשוטות, עמילופלסטים מתחילים להתפתח בשלב מסוים בדרך שונה במקצת. מדענים, לעומת זאת, למדו משהו מעניין: הם הצליחו להשיג את ההפיכה ההדדית של כלורופלסטים מעלי תפוח אדמה לעמילופלסטים (ולהיפך). דוגמה קנונית הידועה לכל תלמיד בית ספר היא שפקעות תפוחי אדמה הופכים לירוקים באור.
אנו יודעים כי בתהליך הבשלת הפירותעגבנייה, תפוחים ועוד כמה צמחים (ובעלי עצים, עשבים ושיחים בסתיו) יש תהליך של "השפלה" כאשר כלורופלסטים בתא צמח הופכים לכרומופלסטים. אברונים אלה מכילים פיגמנטים צבעוניים, קרוטנואידים.
שינוי זה נובע מכך שבמקרים מסוימיםבתנאים, מתרחש הרס מוחלט של תילקואידים, ולאחר מכן האברון רוכש ארגון פנימי אחר. זה המקום בו אנו חוזרים לנושא שעליו התחלנו לדון כבר בתחילת המאמר: השפעת הגרעין על התפתחות כלורופלסטים. זה, באמצעות חלבונים מיוחדים המסונתזים בציטופלסמה של תאים, שיוזם את תהליך ארגון מחדש של האורגנואיד.
לאחר שדיברנו על סוגיות המקור וההתפתחות של כלורופלסטים, צריך להתעכב על המבנה שלהם ביתר פירוט. יתר על כן, זה מעניין מאוד וראוי לדיון נפרד.
המבנה הבסיסי של כלורופלסטים מורכב משנייםממברנות ליפופרוטאין, פנימיות וחיצוניות. העובי של כל אחד מהם הוא כ- 7 ננומטר, המרחק ביניהם הוא 20-30 ננומטר. כמו במקרה של פלסטידים אחרים, השכבה הפנימית יוצרת מבנים מיוחדים הבולטים לאורגנואיד. בכלורופלסטים בוגרים יש שני סוגים של קרומים "מפותלים" כאלה בבת אחת. הראשונים יוצרים את הלמלות הסטרומליות, האחרות - הממברנות התילקואידיות.
יש לציין שיש קשר ברור,שיש לו קרום כלורופלסט עם תצורות דומות בתוך האורגנואיד. העובדה היא שחלק מקפליו יכולים להתרחב מקיר אחד למשנהו (כמו במיטוכונדריה). כך שהלמלות יכולות ליצור סוג של "תיק" או רשת ענפה. עם זאת, לרוב מבנים אלו ממוקמים מקבילים זה לזה ואינם קשורים בשום דרך זה לזה.

המספר הכולל של דגנים הכלולים בכלורופלסטים של צמחים גבוהים יותר, זה יכול להגיע 40-60. כל תילקואיד מחובר כל כך חזק לשני, עד שהקרומים החיצוניים שלהם יוצרים מישור יחיד. עובי השכבה בצומת יכול להיות עד 2 ננומטר. שים לב שמבנים כאלה, שנוצרים על ידי תילאקואידים ולמלות סמוכים, אינם נדירים.
במקומות של קשר שלהם יש גם שכבה,לפעמים מגיעים לאותו 2 ננומטר. לפיכך, כלורופלסטים (שמבנהם ותפקידיהם מורכבים מאוד) אינם מבנה מונוליטי אחד, אלא מעין "מצב בתוך מדינה". בהיבטים מסוימים, מבנה האברונים הללו מורכב לא פחות מכל המבנה הסלולרי!
הגרגירים מתקשרים זה עם זה בדיוק בעזרתלמלה. אך חללי התילקואידים, היוצרים ערימות, סגורים תמיד ואינם מתקשרים עם המרחב הבין-ממשי בשום צורה שהיא. כפי שאתה יכול לראות, מבנה הכלורופלסטים מורכב למדי.
מה יכול להכיל בסטרומה של כולםכלורופלסט? יש מולקולות DNA בודדות וריבוזומים רבים. ב amyloplasts, זה סטרומה כי הם דגני עמילן מופקדים. לפיכך, בכרומופלסטים יש פיגמנטים צבעוניים שם. כמובן, ישנם פיגמנטים שונים של כלורופלסט, אך הנפוץ ביותר הוא כלורופיל. הוא מחולק למספר סוגים בו זמנית:
יש אצות אדומות וחומותכלורופלסטים לא כל כך לעתים רחוקות יכולים להיות סוגים שונים לחלוטין של צבעים אורגניים. אצות מסוימות מכילות בדרך כלל כמעט את כל פיגמנטים הכלורופלסטים הקיימים.
כמובן שתפקידם העיקרי הואהמרת אנרגיית אור לרכיבים אורגניים. הפוטוסינתזה עצמה מתרחשת בגרגרים בהשתתפות ישירה של כלורופיל. הוא סופג את אנרגיית אור השמש והופך אותה לאנרגיה של אלקטרונים נרגשים. האחרון, שיש לו עודף מההיצע שלו, מוותר על אנרגיה עודפת, שמשמשת לפירוק מים וסינתזת ATP. כאשר מים מתפרקים נוצרים חמצן ומימן. הראשון, כפי שכבר כתבנו לעיל, הוא תוצר לוואי ומשתחרר לחלל שמסביב, ומימן נקשר לחלבון מיוחד, פרדוקסין.

ה- ATP המתקבל הוא חשוב ביותר מכיוון שהואה"מצבר "העיקרי של האנרגיה, ההולך לצרכים שונים של התא. NADPH-H2 מכיל חומר הפחתה, מימן, ותרכובת זו יכולה לתת אותו בקלות במידת הצורך. במילים פשוטות, זהו חומר הפחתה כימי יעיל: בתהליך הפוטוסינתזה מתרחשות תגובות רבות, שפשוט אינן יכולות להמשיך בלעדיה.
לאחר מכן, אנזימים של כלורופלסט נכנסים לשחק,הפועלים בחושך ומעבר: המימן מחומר ההפחתה והאנרגיה של ATP משמשים את הכלורופלסט בכדי להתחיל בסינתזה של מספר חומרים אורגניים. מאחר שהפוטוסינתזה מתרחשת בתנאי אור טובים, התרכובות המצטברות בחושך משמשות לצרכי הצמחים עצמם.
אתה יכול לציין בצדק כי תהליך זה דומה באופן מחשיד לנשימה בהיבטים מסוימים. במה שונה הפוטוסינתזה ממנה? הטבלה תעזור לך להבין את הנושא הזה.
נקודות השוואה | פוטוסינתזה | נשימה |
מתי קורה | רק ביום, באור השמש | בכל עת |
לאן זה לוקח | תאים המכילים כלורופיל | כל התאים החיים |
חמצן | הקצבה | קליטה |
CO2 | קליטה | הקצבה |
חומר אורגני | סינתזה, מחשוף חלקי | פיצול בלבד |
אנרגיה | ספג | בולט |
כך שונה הפוטוסינתזה מהנשימה. הטבלה מראה בבירור את ההבדלים העיקריים ביניהם.
רוב התגובות הנוספות מתרחשות כאן.אותו דבר, בסטרומה של הכלורופלסט. הדרך הנוספת של החומרים המסונתזים שונה. לכן, סוכרים פשוטים עוזבים מיד את האורגנואיד, ומצטברים בחלקים אחרים של התא בצורה של פוליסכרידים, בעיקר עמילן. בכלורופלסטים נוצרים גם שקיעת שומנים וגם הצטברות מקדימה של קודמיהם, המופרשים אזורים אחרים בתא.
צריך להבין בבירור שכל תגובות הסינתזהדורשים כמויות אדירות של אנרגיה. המקור היחיד שלה הוא כל אותה פוטוסינתזה. זהו תהליך שלעתים קרובות דורש כל כך הרבה אנרגיה שיש להשיג אותו באמצעות השמדת החומרים שנוצרו כתוצאה מהסינתזה הקודמת! לפיכך, מרבית האנרגיה המתקבלת במהלך הוצאתה לביצוע תגובות כימיות רבות בתוך תא הצמח עצמו.

מקובל כי אברונים תאיים, כוללכולל כלורופלסטים (המבנה והתפקודים שתארנו בפירוט) ממוקמים במקום אחד בלבד. זה לא נכון. כלורופלסטים יכולים לנוע בתא. לכן, בתאורה חלשה, הם נוטים לתפוס עמדה ליד הצד המואר ביותר של התא, בתנאים של תאורה בינונית ונמוכה הם יכולים לבחור כמה עמדות ביניים בהן ניתן "לתפוס" את מרבית אור השמש. תופעה זו נקראת "פוטוטקסיס".
כמו המיטוכונדריה, כלורופלסטים הםאברונים די אוטונומיים. יש להם ריבוזומים משלהם, הם מסנתזים מספר חלבונים מאוד ספציפיים המשמשים רק אותם. ישנם אפילו מתחמי אנזים ספציפיים שבמהלך עבודתם מיוצרים שומנים מיוחדים הנדרשים לבניית ממברנות הלמלה. כבר דיברנו על מקורם הפרוקריוטי של אברונים אלה, אך יש להוסיף כי ישנם מדענים הרואים בכלורופלסטים צאצאים ותיקים של כמה אורגניזמים טפיליים, שהפכו תחילה לסימביונטים, ואז הפכו לחלוטין לחלק בלתי נפרד מהתא.
עבור צמחים זה ברור - זו סינתזה של אנרגיה וחומרים המשמשים את תאי הצמח. אך פוטוסינתזה היא תהליך המבטיח הצטברות מתמדת של חומרים אורגניים בכל כדור הארץ. כלורופלסטים יכולים לסנתז כמות עצומה של התרכובות המורכבות ביותר מולקולריות גבוהות מפחמן דו חמצני, מים ואור שמש. יכולת זו אופיינית רק להם, ואדם עדיין רחוק מלחזור על תהליך זה בתנאים מלאכותיים.

אנו מקווים שלמדת ממאמר זה על מהו כלורופלסט ומה תפקידו באורגניזם הצמחי.


