


Шпинделът на разделянето е временна структураобразуват се по време на процесите на митоза и мейоза и осигуряват сегрегацията на хромозомите и клетъчното делене. Тя е биполярна: микротубулната система, образувана в пространството между полюсите, наподобява форма на вретено. В областта на центромера, микротубулите на вретено се свързват с хромозомите кинетохори. На тях хромозомите се движат към полюсите.

Шпинделът на разделянето се състои от три основниструктурни елементи: микротубули, деление на полюси и хромозоми. Полюсите на разделяне при животните се организират с помощта на центрозоми, които съдържат центриоли. В случай на отсъствие на центрозоми (при растения и в ооцити при някои животински видове), вретеното има широки полюси и се нарича ацентрозомално. Друга структура е включена в образуването на вретено-моторни протеини. Те принадлежат на dyneinam и kinesinam.
Веретено деления - это биполярная структура.На двата полюса са разположени центрозоми - органели, които са центровете на микротубулната организация. В структурата на центрозома има две центриоли, които са заобиколени от много различни протеини. Между полюсите са разположени кондензирани хромозоми, които имат формата на две хроматиди, прикрепени към центромерно място. В областта на центромера има кинетохори, към които са прикрепени микротубули.
Тъй като шпинделът за разделяне е структура,отговорен за клетъчното делене, началото на неговото сглобяване се среща в профаза. В растенията и в яйцеклетките, в отсъствието на центрозоми, обвивката на ядрото служи като център на микротубулната организация. Микротубулите се доближават до ядрената обвивка и в края на профазата завършват ориентацията им и се образува "профазен шпиндел" - оста на бъдещото разделение на вретеното.
Поради факта, че в животински клеткиЦентрозома действа като център на организацията, като началото на образуването на вретеното на деление е дивергенцията на двата центрозома по време на профазния период. Това е възможно поради двигателните протеини dyneinam: те са прикрепени към външната повърхност на ядрото, както и от вътрешната страна на клетъчната мембрана. Групата от диенини, прикрепени към мембраната, се свързва с астралните микротубули и те започват да се движат към минус края, поради което центрозомите се разреждат в противоположни части на клетъчната мембрана.
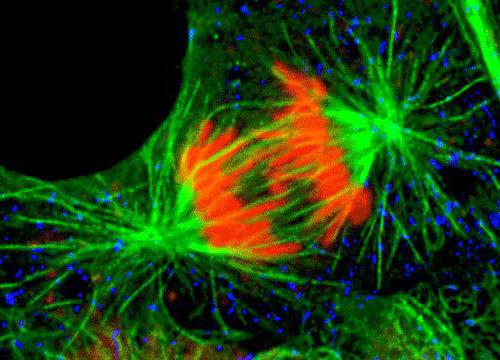
Окончательное формирование веретена деления настъпва на етапа на прометафаза, след изчезването на мембраната на ядрото, тя става пълноценна, тъй като след това центрозома и микротубулите могат да достигнат компонентите на вретеното.
Въпреки това, има едно изключение: при маточните дрожди, образуването на вретеното на делене се случва вътре в ядрото.
Образуването на влакна вретено и технитеориентацията е невъзможна без два процеса: организирането на микротубулите около хромозомите и привързаността им един към друг в противоположните полюси на разделението. Много от елементите, необходими за окончателното формиране на разделящото вретено, включително хромозоми и моторни протеини, са вътре в клетъчното ядро и микротубули и, ако е животинска клетка, центрозомите се съдържат в цитоплазмата, т.е. компонентите са изолирани един от друг. Ето защо образуването на вретеното завършва едва след изчезването на ядрената обвивка.

При формирането на делителното вретено се включва протеин, иима и много други структури и този процес е добре проучен в животински клетки. По време на профазния период микротубулите образуват звездообразна структура около центрозома, която се отклонява в радиалната посока. След като мембраната на ядрото се разруши, динамично нестабилните микротубули започват активно да изследват тази област и кинохорите на хромозомите могат да бъдат фиксирани върху тях. Някои от хромозомите веднага се появяват в противоположни полюси, а останалите се свързват първо с микротубули на един от полюсите и едва след това започват да се движат към желания полюс. Когато процесът приключи, хромозомите, вече асоциирани с полюс, започват да се прикрепват към микротубулите от противоположния полюс с кинетохори, като по този начин от десет до четиридесет тръби са прикрепени към кинетохорите по време на метафазния процес. Тази формация се нарича кинетохорна греда. Постепенно, всяка от хромозомите се свързва с противоположния полюс и те образуват метафазна плоча в централната част на разделителното вретено.

Има и друг сценарий, в койтооформено вретено. Това е възможно за клетки с центрозоми и за клетки, в които те липсват. Гама-тубулинов пръстен е включен в процеса, поради което около хромозомите възниква нуклеация на къси микротубули. Тубулите са прикрепени към кинетохорите с плюс-край, след което започва полимеризацията на микротубулите, т.е. контролирания растеж. Отрицателните краища "се сливат" и остават в полюсите на деление поради моторните протеини. Ако една двойка центрозома участва в образуването на разделящото вретено, това улеснява връзката на микротубулите, но процесът е възможен без тях.

Ясно отделяне на хромозомите между две клеткиобразувани по време на деленето, могат да възникнат само ако двойките хроматиди с техните кинетохори свързват различни полюси. Биполярното несъответствие на хроматидите се нарича амфитепично, но има и други варианти, които се появяват по време на сглобяването на разделящото вретено. Това е монотепично (един кинетохор е свързан със същия полюс) и синтетичен (двете кинетохори на хромозомата са свързани с един полюс). В меротепичния, кинетохорът се улавя веднага от два полюса. Само нормално биполярно свързване, което се дължи на силите на напрежение от полюсите, е стабилно, други методи на свързване са нестабилни и обратими, но са възможни поради разположението на кинетохорите.


