


Divisjonsspindelen er en midlertidig struktur,dannet under prosessene med mitose og meiose, og gir kromosomsegregasjon og celledeling. Det er bipolært: mikrotubulesystemet dannet i rommet mellom polene ligner en spindel i form. I sentromerregionen blir spindelmikro-rørkuler sammenføyning med kinetokorene i kromosomet. På dem beveger kromosomene seg til polene.

Spindelen til divisjon består av tre hovedstrukturelle elementer: mikrotubuler, fisjoneringspoler og kromosomer. Delingspolene hos dyr er organisert ved hjelp av sentrosomer, som inneholder sentrioler. I mangel av et sentrosom (i planter, og i oocytter hos noen dyrearter), har spindelen brede poler og kalles acentrosomal. I dannelsen av spindelen er en annen struktur involvert - motoriske proteiner. De tilhører dyneiner og kinesiner.
Delingsspindelen er en bipolar struktur.Ved begge polene er sentrosomer - organeller, som er sentrene for mikro-rørorganisering. I sentrosomstrukturen skilles to sentrioler, omgitt av mange forskjellige proteiner. Kondenserte kromosomer i form av to kromatider bundet til et sentromersted er plassert mellom polene. I området av sentromerene er det kinetokorer, som mikrotubuli knytter seg til.
Siden divisjonsspindelen er en struktur,som er ansvarlig for celledeling, begynner begynnelsen av samlingen i profase. I planter og i oocytter, i fravær av sentrosomer, er kjernemembranen sentrum for mikro-rørorganiseringen. Mikrotubuljer nærmer seg kjernekonvolutten, og på slutten av profesen slutter deres orientering og en "profasespindel" dannes - aksen til den fremtidige splittelsespindelen.
Med tanke på det faktum at nettopp i dyrecellersentrosomet fungerer som sentrum for organisasjonen, begynnelsen på dannelsen av divisjonsspindelen er avviket mellom to sentrosomer under profesen. Dette er mulig takket være motoriske proteiner dyneiner: de fester seg til ytre overflate av kjernen, så vel som på innsiden av cellemembranen. Gruppen av dyneiner festet til membranen er koblet til de astrale mikrotubuli og de begynner å bevege seg mot minusenden, på grunn av hvilken sentrosomet blir fortynnet i motsatte deler av cellemembranen.
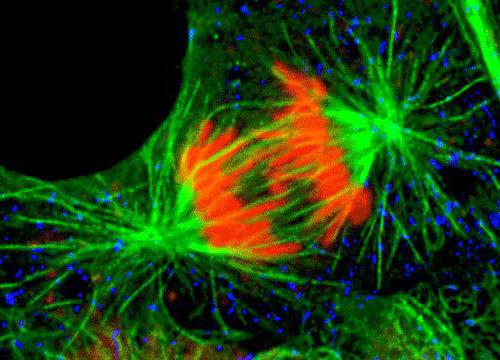
Endelig spindeldannelseoppstår på prometafase-stadiet, etter at kjernemembranen forsvinner, blir den fullverdig, fordi det er etter dette sentrosomene og mikrotubuli kan få tilgang til komponentene i spindelen.
Imidlertid er det ett unntak: i spirende gjær oppstår dannelsen av en fisjonspindel inne i kjernen.
Dannelse av spindeltråder og deresorientering er umulig uten to prosesser: organiseringen av mikrotubuli rundt kromosomene og deres tilknytning til hverandre ved motsatte fisjon poler. Mange elementer som er nødvendige for den endelige dannelsen av divisjonsspindelen, inkludert kromosomer og motoriske proteiner, er plassert inne i cellekjernen, og mikrotubuli, og hvis det er en dyrecelle, er sentrosomer inneholdt i cytoplasma, det vil si at komponentene er isolert fra hverandre. Det er derfor dannelsen av spindelen slutter først etter at kjerneskallet forsvant.

Protein er involvert i dannelsen av fisjonspindelen, ogogså mange andre strukturer, og i dyreceller er denne prosessen godt studert. I løpet av profase-perioden danner mikrotubuli en stjerneformet struktur rundt sentrosomet, som avviker i radiell retning. Etter at kjernemembranen er ødelagt, begynner dynamisk ustabile mikrotubuler aktivt å undersøke dette området, og kinetokorer av kromosomer kan festes på dem. En del av kromosomene vises umiddelbart ved motsatte poler, mens resten først er assosiert med mikrotubuli av en av polene, og først da begynner de å bevege seg mot ønsket pol. Når prosessen er fullført, begynner kromosomene som allerede er koblet til en pol å bli festet av kinetokorer til mikrotubuli fra den motsatte polen, og dermed blir ti til førti rør festet til kinetokorene under metafaseprosessen. Denne formasjonen kalles en kinetokore bjelke. Gradvis blir hvert av kromosomene assosiert med den motsatte polen, og de danner en metafaseplate i den sentrale delen av fisjonspindelen.

Det er et annet scenario derå danne en spindel av divisjon. Dette er mulig både for celler med sentrosomer og for celler der de er fraværende. Gamma-tubulin ringkomplekset er involvert i prosessen, på grunn av hvilken det er kjernevirkning av korte mikrotubuli rundt kromosomene. Rørene festes til kinetokorene med plussenden, hvoretter polymeriseringen av mikrotubuli begynner, det vil si regulert vekst. Minusen ender "fusjonere" og forblir ved delingspolene på grunn av motoriske proteiner. Hvis et sentrosompar er involvert i dannelsen av fisjoneringsspindelen, letter dette forbindelsen av mikrotubuli, men prosessen er mulig uten dem.

Tydelig separasjon av kromosomer mellom to celler,dannet under fisjon, kan bare skje hvis sammenkoblede kromatider, med deres kinetokorer, blir med i forskjellige poler. Det bipolare avviket mellom kromatider kalles amfittepisk, men det er andre alternativer som oppstår under delingen av spindelen. Dette er monotepisk (en kinetokore blir en pol) og syntepisk (begge kinetokore kromosomer kobles til en pol). I tilfelle av en merotepisk blir en kinetokore fanget på en gang av to poler. Stabil er bare den vanlige, bipolare bindingen, som oppstår på grunn av trekkreftene fra polene, andre bindingsmetoder er ustabile og reversible, men mulig på grunn av kinetokorens beliggenhet.


